|
|
 |
THE BRAIN STEM
 General anatomical considerations: General anatomical considerations:
The diencephalon along with the midbrain, pons, and medulla
oblongata comprise the brainstem. A clear understanding of the
importance of this area of the CNS requires that to be familiar
with both its external and internal features. In addition to
performing many vitally important regulatory functions
(respiratory and cardiovascular), the brainstem also serves as a
central point of relay between the cerebrum, the cerebellum, and
the receptors and effectors of the cranial and spinal nerves.
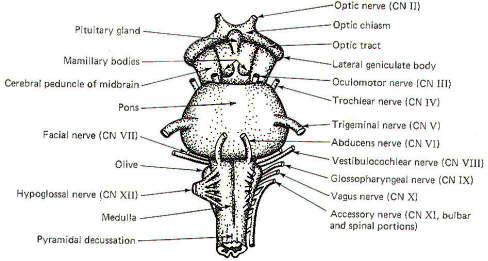
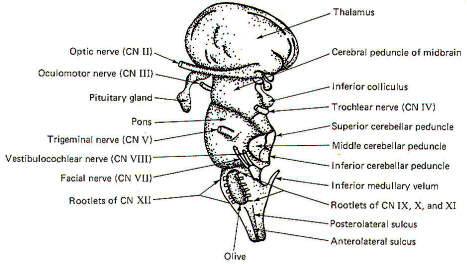
EXTERNAL MORPHOLOGY
The prominent external features of the brain stem are
illustrated in Fig-1,2,3. The cerebrum and cerebellum have been removed in
each drawing in order to afford an unobstructed view of the
brainstem from anterior, posterior, and lateral perspectives.
The Midbrain
The most prominent features of the anterior and lateral midbrain
are the cerebral peduncles. These broad bundles of descending
fibers from the cerebrum converge to form a V on the anterior
surface, bounded above by the optic chiasm and below by the
superior border of the pons. The mammillary bodies and the
pituitary gland are framed by the two peduncles. Four prominent
enlargements, the corpora quadrigemina, can be seen on the
posterior surface of the midbrain. The quadrigemina (four
bodies) include two superior colliculi and two inferior
colliculi. The trochlear nerves (IV) emerge from the posterior
surface of the midbrain just below the inferior colliculi,
wrapping around the cerebral peduncles to appear anterolaterally
at the superior border of the pons. The oculomotor nerves (III)
also originate in the midbrain, emerging anteriorly at the
superior border of the pons.
The Pons
The pons is a distinctively prominent feature of the brainstem.
It appears as a broad band of transversely running fibers when
viewed anteriorly and from the side. The fibers extend into the
cerebellum behind and appear to be holding it to the brainstem.
Those which wrap laterally to the cerebellum form the middle
cerebellar peduncles.
The pons is bounded superiorly by the midbrain and inferiorly by
the medulla oblongata. The trigeminal nerves (V) are prominent
lateral projections. The abducens nerves (VI) originate in the
pons and emerge close together at the anterior inferior border
of the pons. The facial nerves (VII), originating in the pons,
and the vestibulocochlear nerves (VIII), originating in the
pontomedullary area, emerge at the pontomedullary border.
 |
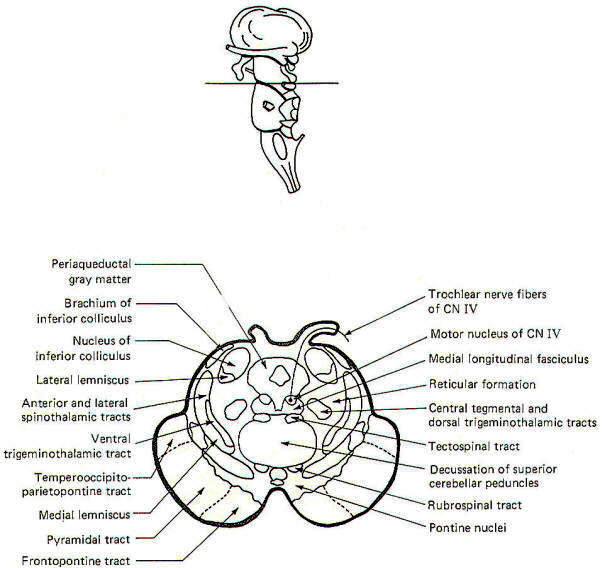 |
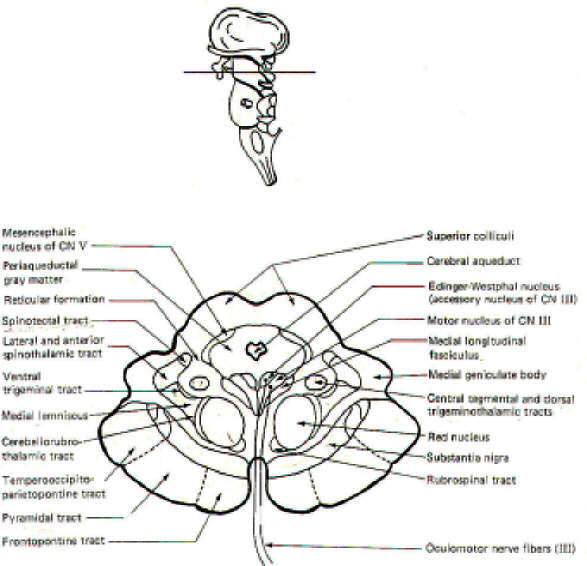
| Fig-4: The mesencephalon -upper
section. |
Fig-5: The mesencephalon- lower
section. |
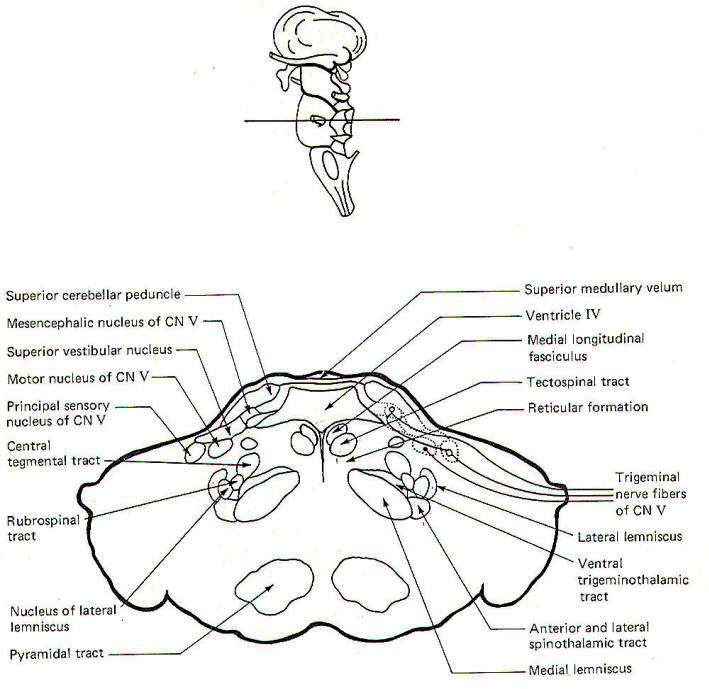 |
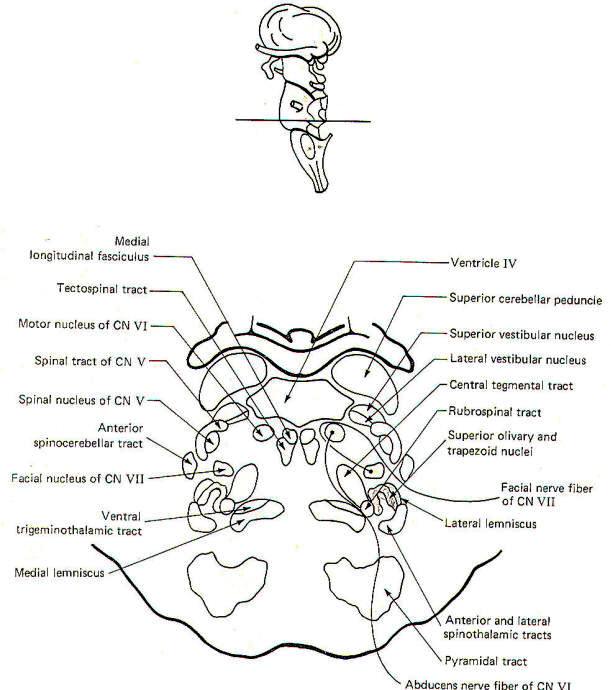 |
| Fig-6: The pons |
Fig-7: The pons |
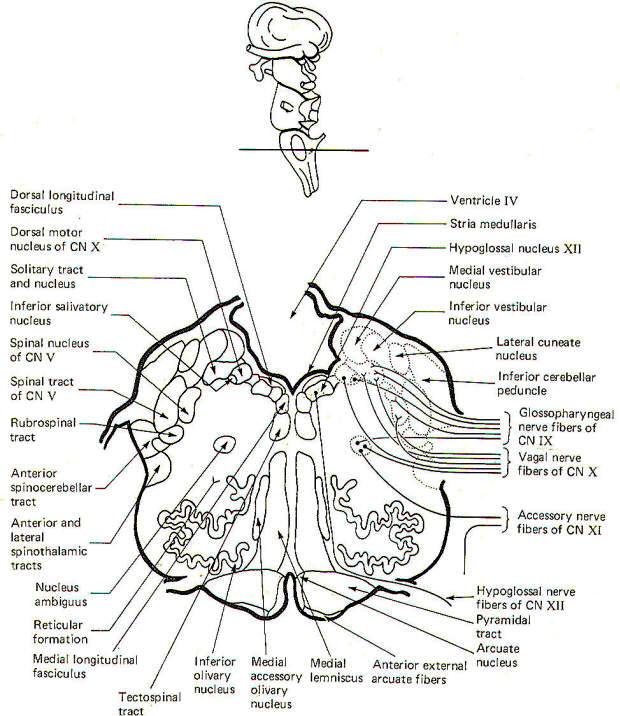
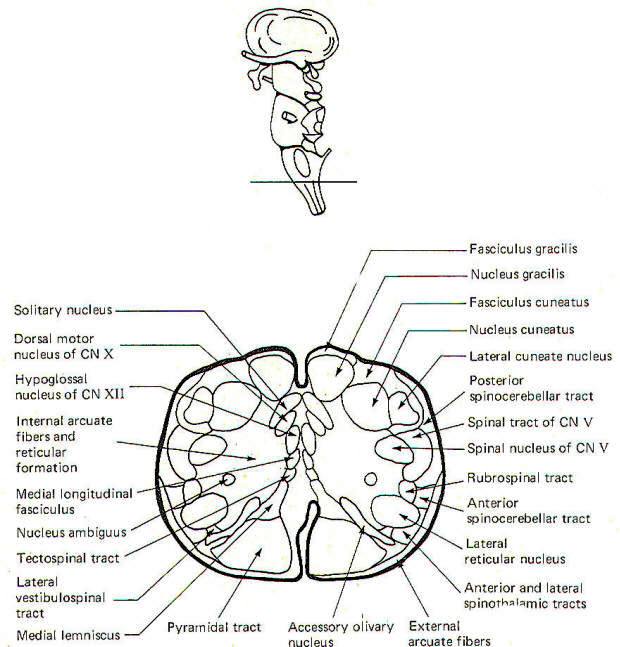
The Medulla Oblongata
The most prominent anterior features of the medulla oblongata
are the medullary pyramids. They appear on the anterior surface
as two vertically running rounded eminences which emerge from
under the pons to become continuous with the spinal cord below.
In the lowest portion of the anterior medulla, descending
corticospinal (pyramidal) tract fibers cross over in the
pyramidal decussation. The corticospinal tracts are often called
pyramidal tracts because of the unique pyramidal shape they give
to the anterior medulla as they descend into the spinal cord.
The olive is a lateral feature of the medulla. Emerging from the
lateral medulla posterior to the olive in descending order are
the glossopharyngeal nerves (IX), the vagus nerves (X), and the
bulbar accessory nerves (XI). The hypoglossal nerves (XII)
emerge from the lateral medulla anterior to the olive.
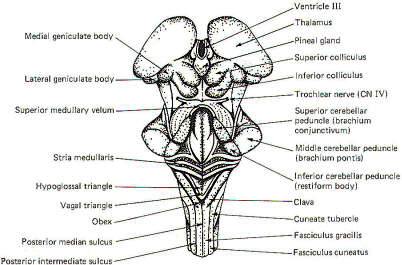
Three sulci are visible in posterior view, a single posterior
median sulcus and two laterally placed posterior intermediate
sulci. Two rounded eminences, the gracile tubercle (clava) and
the cuneate tubercle are observed on either side of the
posterior median sulcus. The fasciculus gracilis leads to the
former while the fasciculus cuneatus leads to the latter. The
posterior intermediate sulcus separates the fasciculus gracilis
and gracile tubercle from the fasciculus cuneatus and cuneate
tubercle on either side.
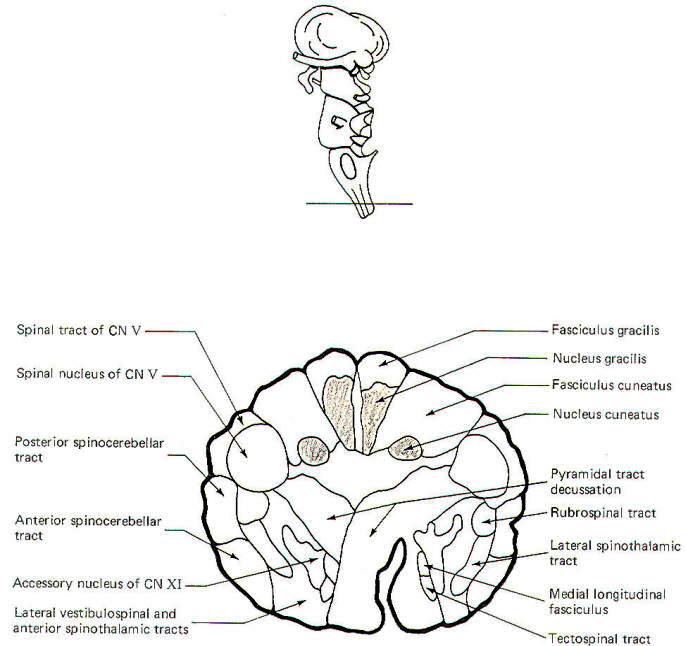
CROSS-SECTIONAL ANATOMY OF THE BRAINSTEM
As pathways ascend and descend through the brainstem they often
undergo shifts in position which can only be seen by a careful
examination of cross-sectional anatomy. This is verified by
close examination of the eight representative sections
schematically illustrated in Figs-4 through 11. There is no
real shortcut or alternative to "learning" these cross sections.
Indeed, the function of the brainstem as a relay center between
the cerebrum above, the cerebellum behind, and the spinal cord
below is easier to visualize.
As an academic exercise, it is useful to follow the course of
pathways through the brainstem. By doing this it is possible to
observe how the tracts change in relative position and size as
they descend through the stem. For example, the corticospinal
tracts enter the brainstem in the middle third of the basis
pedunculi (ventral portion) of the cerebral peduncles where they
are widely separated from each other. As they descend through
the pons they move to a deeper position away from the surface.
However, upon entering the medulla they begin to converge and
once again move to the surface, giving rise to the medullary
pyramids. Bundles of crossing fibers of these tracts can be
observed in the pyramidal decussation in the lower medulla.
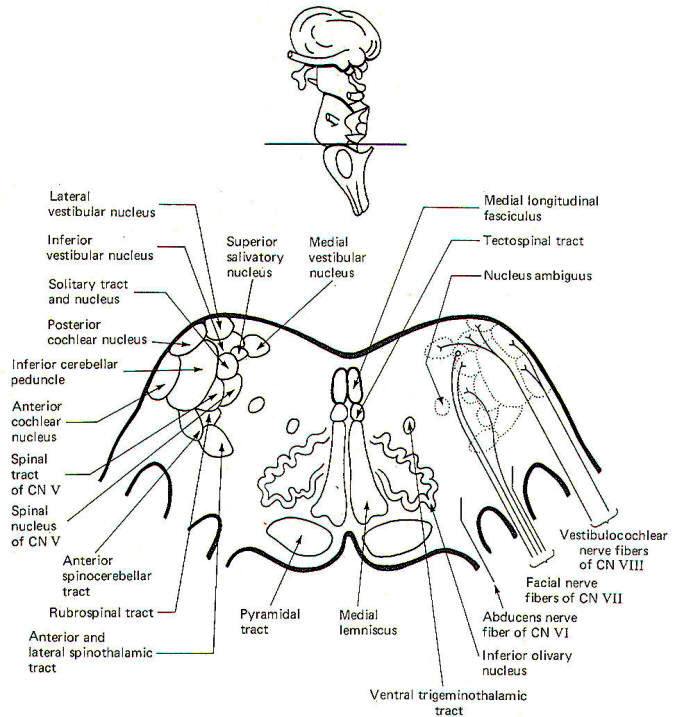 |
 |
| Fig-8: Cross section at the
pontomedullary junction. |
Fig-9: |
 |
 |
| Fig-10 |
Fig-11 |
 |
 |
| Fig-12 |
Fig-13 |
CRANIAL NERVES AND BRAINSTEM NUCLEI
Cranial Nerve Fiber Classification
Cranial nerve fibers are classified as general or special,
somatic or visceral, and afferent or efferent.
Special fibers are those which innervate the special sense
organs associated with hearing, seeing, smelling, and tasting.
In addition, they innervate the vestibular apparatus and those
skeletal muscles derived from the mesoderm of the branchial
arches. This latter group includes the muscles of facial
expression and mastication as well as laryngeal and pharyngeal
muscles. Also included are the sternomastoid and trapezius
muscles. All other cranial nerve fibers are classified as
general.
Fibers are further designated somatic or visceral. Somatic
fibers innervate those skeletal muscles derived from mesodermal
somites as well as innervating structures of ectodermal origin.
The latter include the skin, the eye, the vestibular apparatus,
and the inner ear. Exceptions are the olfactory epithelium and
the taste buds. Even though the olfactory epithelium and taste
buds are of ectodermal origin, the cranial nerve fibers
innervating them are classified as visceral because of the close
functional relationship which the senses of smell and taste have
with the truly visceral gastrointestinal tract.
Visceral fibers innervate structures of entodermal origin
including cardiac muscle, smooth muscle, and glands. Also
included here are those skeletal muscles derived from the
mesoderm of the branchial arches. As previously noted, cranial
nerve fibers mediating smell (I) and taste (VII, IX, and X) are
typically included here rather than with the somatic group.
Cranial nerve fibers are also classified as afferent or
efferent, depending on the direction of their impulse
conduction. Afferent fibers conduct impulses toward the CNS
while efferent fibers conduct them away.
An oddity in the classification scheme arises from the practice
of classifying all proprioceptors as general somatic regardless
of whether they are associated with somatic or branchial
muscles. This leads to the confusing observation that a muscle
can be innervated by both special visceral efferent and special
somatic afferent fibers at the same time. The muscles of
mastication are an example (Fig-14). The scheme of cranial
nerve fiber classification is presented again in Table-1.
 |
 |
| Fig-14 |
Fig-15 |
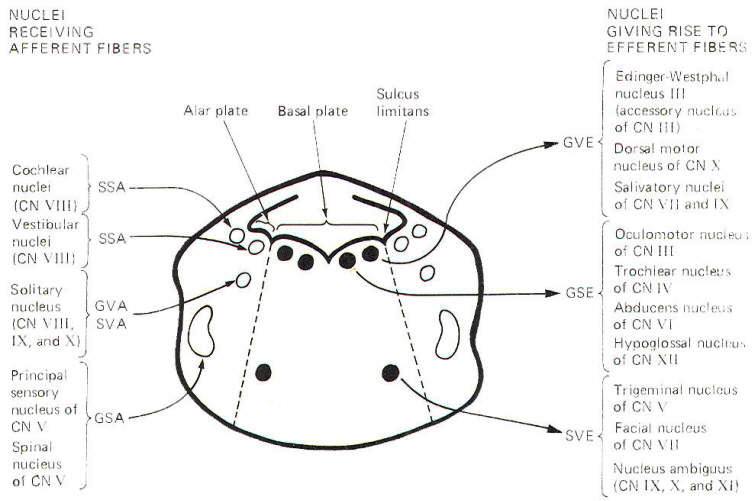
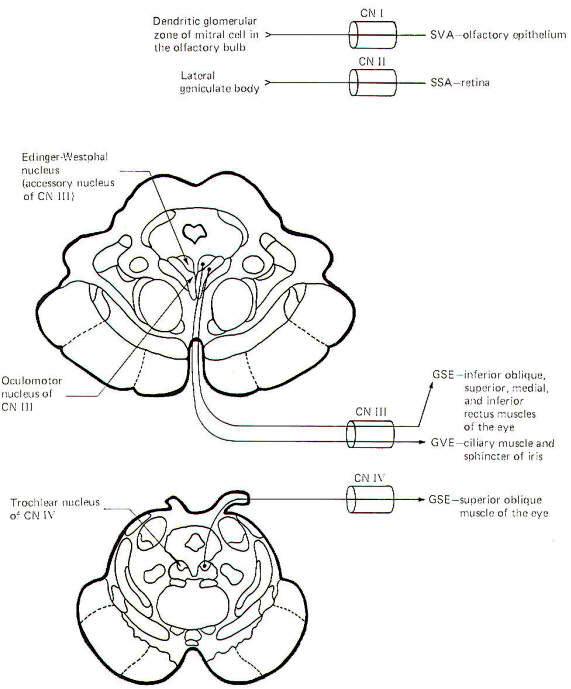
Alar and Basal Nuclei
When the embryonic neural tube closes, a groove remains in each
lateral wall which separates the posterior from the anterior
portions. The former gives rise to the alar lamina or plate,
while the latter forms the basal lamina (Fig. 9-12). Brainstem
sensory nuclei are found in the alar lamina, while motor nuclei
are generally distributed in the basal lamina. Figure 9-12 is a
composite sketch of cranial nerve nuclei as found in the
brainstem from the midbrain to the medulla oblongata. It is not
a sketch of any single brain stem section but instead represents
a construct intended to show the relative positions of the
nuclei with respect to each other in cross section. Notice that
the efferent (motor) nuclei are located in the basal plate,
while the afferent (sensory) nuclei are located in the more
lateral alar plates. The dividing line is the sulcus limitans.
Cranial Nerve Fibers and the Brainstem
It is not too difficult to trace the emergence of each cranial
nerve from the brainstem. A more difficult task is to appreciate
the distinct fiber types present in each cranial nerve. But
unquestionably the most difficult task of all is to trace the
efferent origins and afferent terminations of the cranial nerve
fibers in the brainstem. These relationships are illustrated in
Figs. 9-13 through 9-15.
Olfactory Nerve (I): The fibers of this nerve are SVA. They
carry information pertinent to smell from the olfactory
epithelium to the dendritic glomerular zone of the mitral cells
in the olfactory bulb. Mitral cell fibers then conduct smell
information to the olfactory cortex. Damage to these tracts
causes anosmia (loss of the sense of smell).
Optic Nerve (II): The fibers of this nerve are SSA. They conduct
information concerning vision from the ganglion cell layer of
the retina primarily to the lateral geniculate bodies. Damage to
these fibers causes anopsia (loss of vision).
Oculomotor Nerve (III): The oculomotor nerve contains GVE and
GSE fibers. The GVE fibers originate in the Edinger-Westphal
nucleus (an accessory nucleus of III) in the upper midbrain.
They represent the preganglionic parasymapthetic fibers to the
ciliary ganglion. Postganglionic fibers innervate the ciliary
muscles, which control the thickness of the lens, as well as the
sphincter muscles of the iris, which control pupil size. Damage
to these fibers eliminates the pupillary light reflex and
interferes with accommodation reflexes.
The GSE fibers originate in the oculomotor nucleus in the upper
midbrain. They innervate the inferior oblique as well as the
superior, medial, and inferior rectus muscles of the eye. Damage
to these fibers results in external strabismus and ptosis of the
eyelid.
Trochlear Nerve (IV): The fibers of this nerve are GSE. They
originate in the trochlear nucleus of the lower midbrain. They
innervate the superior oblique muscles of the eye. Damage to
these fibers causes the eyes to look slightly upward.
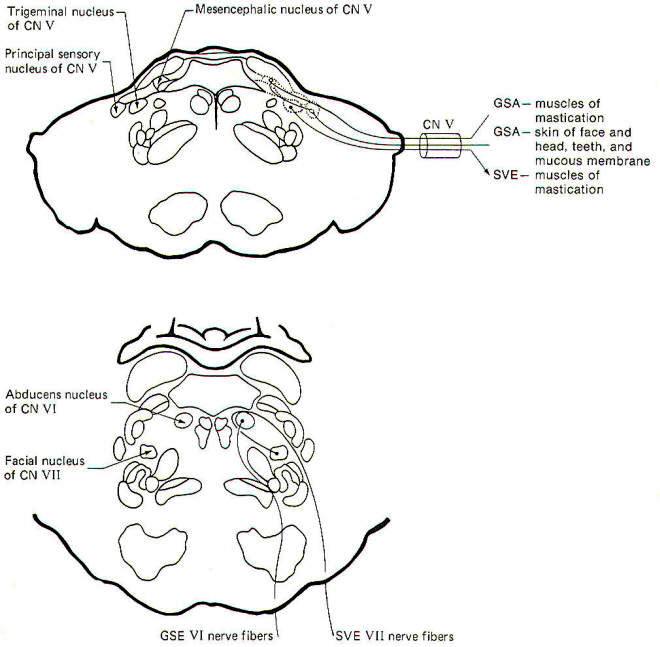
Trigeminal Nerve (V): The trigeminal nerve contains SVE and GSA
fibers. The SVE fibers originate in the trigeminal nucleus
located in the middle pons. They innervate the muscles of
mastication (branchiomeric origin). Damage to these muscles
causes paralysis of the jaws.
GSA fibers fall into two groups, those from proprioceptors and
those from exteroceptors. Proprioceptive fibers have their cell
bodies in the mesencephalic nucleus of V and terminate in the
principal sensory nucleus of V in the pons. Exteroceptive fibers
from the skin of the face and head as well as the teeth and the
mucous membranes conduct information to the principal sensory
nucleus of V. Damage to these fibers causes anaesthesia in the
affected area.
Abducens Nerve (VI): The fibers of this nerve are GSE. They
originate in the abducens nucleus in the lower pons and
innervate the lateral rectus muscle of the eye. Damage to these
fibers causes internal strabismus and double vision.
Facial Nerve (VII): The facial nerve is composed of SVE, GVE,
GSA, and SVA fibers. The SVE fibers originate in the facial
nucleus of the pons and innervate the muscles of facial
expression. Damage to these fibers causes facial paralysis. The
GVE fibers are the preganglionic parasympathetic fibers to the
submaxillary ganglion. They originate in the superior salivatory
nucleus of the pontomedullary region. Postganglionic fibers
innervate the submaxillary and sublingual salivary glands.
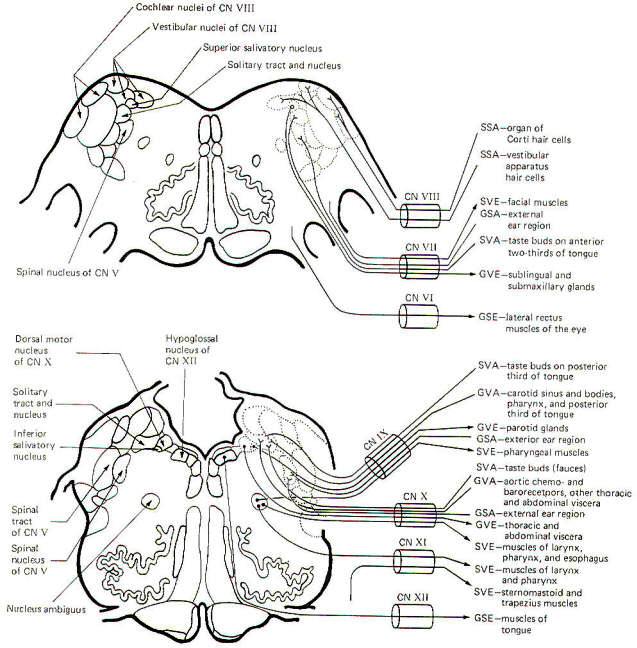
The GSA fibers conduct information from the skin of the external
ear region to the spinal tract and nucleus of V. The SVA fibers
conduct information from the taste buds on the anterior
two-thirds of the tongue to the solitary tract and nucleus.
Vestibulocochlear Nerve (VIII): The fibers of this nerve are
SSA. SSA fibers from the organ of Corti hair cells conduct
auditory information to the cochlear nuclei of the
pontomedullary region. SSA fibers from the vestibular apparatus
hair cells conduct information concerning equilibrium to the
vestibular nuclei in the same general region.
Glossopharyngeal Nerve (IX): The glossopharyngeal nerve is
composed of GVE, SVE, GVA, GSA, and SVA fibers. The GVE fibers
originate in the inferior salivatory nucleus. These are
preganglionic parasympathetic fibers to the otic ganglion.
Postganglionic fibers innervate the parotid salivary glands. The
SVE fibers originate in the nucleus ambiguus and innervate the
pharyngeal muscles (branchiomeric origin). GVA fibers conduct
information from the pharynx and posterior third of the tongue.
These fibers also innervate the carotid sinus baroreceptors and
carotid body chemoreceptors. Signals are conducted to the
solitary tract and nucleus.
The GSA fibers conduct information from the skin of the external
ear region to the spinal tract and nucleus of V. SVA fibers
carry information from the taste buds on the posterior third of
the tongue to the solitary tract and nucleus.
Vagus Nerve (X): The vagus nerve is composed of GVE, SVE, GSA,
GVA, and SVA fibers. The GVE fibers originate in the dorsal
motor nucleus of X and innervate thoracic and abdominal viscera.
These are the parasympathetic fibers of the vagus nerve. The SVE
fibers innervate the muscles of the larynx and pharynx
(branchiomeric origin) and originate in the nucleus ambiguus.
The GSA fibers carry information from the skin of the ear region
to the spinal tract and nucleus of V. GVA fibers conduct signals
from the aortic baroreceptors and chemoreceptors as well as
other thoracic and abdominal viscera to the solitary tract and
nucleus. Taste cells in the fauces send signals over SVA fibers
to the solitary tract and nucleus.
Accessory Nerve (XI): The fibers of the accessory nerve are SVE.
There are two components to this nerve, a bulbar component
arising from nuclei within the brain stem and a spinal component
arising from nuclei in upper cervical levels of the spinal cord.
The SVE fibers which arise from the nucleus ambiguus of the
medulla innervate the muscles of the larynx and pharynx
(branchiomeric origin). SVE fibers arising in the spinal
accessory nucleus in the upper cervical levels of the cord
innervate the sternomastoid and trapezius muscles (also of
branchiomeric origin).
Hypoglossal Nerve (XII): The fibers of this nerve are GSE. They
originate in the hypoglossal nucleus of the medulla and
innervate the muscles of the tongue.
|
|
|
|
|
Prof. Munir Elias
Our brain is a mystery and to understand it, you
need to be a neurosurgeon, neuroanatomist and neurophysiologist.
neurosurgery.tv

Please visit this site, where daily neurosurgical activities are going
on.

Inomed ISIS IOM System
|
|
|
|